
The reading for this session is linked from the course index page.
We'll start the face-to-face session by covering any questions that have arisen. Please propose and vote for questions below. We'll cover the questions with the most votes in class.
Session 1: Introduction
This course aims to develop the critical analysis and communication skills that characterize effective thinkers.
By visiting the forefront of the discipline, we'll get to grips with the open palaeontological controversies that are exercising current researchers, and enter the frontier territory where there are no "right answers"!
By the end of this term, you and your group will construct a "microsite" that digs into a current palaeontological controversy, which you will summarize orally to your peers.
You will then be tasked with writing an essay that makes a convincing argument based on one of your peers' microsites, convincing a reader of your chosen perspective.
As such, your site must provide its audience with a complete introduction to the subject, so that students interested in tacking the controversy can appreciate the evidence behind the key arguments.
To equip you to engage with the primary literature in a critical fashion, we will start the term by addressing two current controversies as a class. For each topic, we'll start with an informal "primer" seminar introducing key background concepts. We'll then split into groups to discuss a key paper at the centre of the controversy. Your group will then find and read further papers to prepare to debate one side of the argument the following week.
The adjacent figure is a schematic representation of relationships between the deuterostomes – our own subdivision of the animal kingdom.
You may not be familiar with all of these phyla. The questions below will help you to learn how to read and navigate this phylogenetic tree.
More on chordate relationships: Satoh et al. 2014 Chordate evolution and the three-phylum system. Proc. R. Soc. B 281: 20141729.

|
| Silhouettes: Phylopic contributors |
A clade (also termed a monophyletic group) is a set of organisms comprising a single common ancestor and all of its descendants.
Despite their name, hemichordates are more closely related to echinoderms than to chordates.
The most recent common ancestor of these four phyla corresponds to the deuterostome common ancestor – whose descendants also include echinoderms.
This clade, termed "Ambulacraria", is characterized by feeding appendages and a distinctive larval morphology termed the dipleura larva.
Locate the most recent common ancestor of these three taxa – starting at each taxon in turn, trace a path towards the root of the tree. The most recent common ancestor is the first point at which all three paths meet.
Now trace the path from this point to the bottom of the tree. The six lines you cross on this path will correspond to characters present in chordates.
This cladogram only displays the relationships between taxa. The upwards direction denotes a temporal sequence, but is not scaled to time (or any other measure of evolutionary change).
So whilst we can say that the pharynx evolved before the tunic (as certain ancestors of urochordates exhibited a pharynx but not a tunic), we can't tell from the tree whether these events occurred before or after the origin of stereom.
Locate the point on the tree at which the pharynx evolved. Which three phyla are descended from this individual?
Locate the point on the tree at which the brain evolved. Which two phyla are descended from this individual?
Where on the tree is the most recent common ancestor of cephalochordates and vertebrates?
This common ancestor was itself a chordate (the first crown-group chordate), but was no more a cephalochordate than a vertebrate. Both lineages have since undergone ~520 million years of evolution since being descended from this shared common ancestor, and these two independent evolutionary histories are what made each group what they are today.
The tree doesn't contain any absolute time information. We can say that the cephalochordate lineage diverged from the vertebrate lineage before vertebrates diverged from urochordates – but notice that this is a different question. We don't know what other lineages might have diverged from the cephalochordates before the group attained its modern identity.
As the age of a crown group is defined by the age of its most recent common ancestor, we'd need to date the divergence of all living cephalochordate lineages in order to establish the age of the crown-group phylum.
Starting at the urochordates, trace a path to the first node (branching point). What other taxa are descended from this common ancestor?
Starting at the hemichordates, trace a path to the first node (branching point). All echinoderms are descended from this common ancestor, so are equally closely related – just as a grandmother is equally closely related to all of her grandchildren.
Remember that you can rotate any node in the tree without changing the relationship information it contains: mirroring the position of starfish and crinoids would not change their last common ancestor, hence does not affect the relationships portrayed by the tree.
This is a bad question: all the groups are alive today, so each has been evolving for 3 456 million years. Some groups share more of this history than others, depending on the sequence in which the lineages diverged.
Review: Misinterpretations about progress
The important point here is that the shape of the tree depends on which taxa are included; cephalochordates are no 'less evolved' than vertebrates, even if they exhibit a different (and less complex?) set of morphological features.
Groups that are morphologically simple may be secondarily so (as with adult urochordates); groups that are less diverse are not necessarily less sophisticated.




Which group is most closely related to urochordates? To vertebrates?
Remember, the most closely related group is the group that shares a most recent common ancestor. Common ancestors occur at the branching points on a path traced towards the root of the tree.
Gyaltsenglossus is a Cambrian fossil with a three-part body, comprising a proboscis, collar and trunk.
Where on the tree is it most likely to plot?

Unless the three-part body denotes convergent evolution, it belongs in the Hemichordate total group. The information available is insufficient to determine whether it belongs in the crown or stem group.
Banffia is another enigmatic Cambrian fossil from the Burgess Shale; it has a feeding appendage, but lacks a stereom or three-part body.
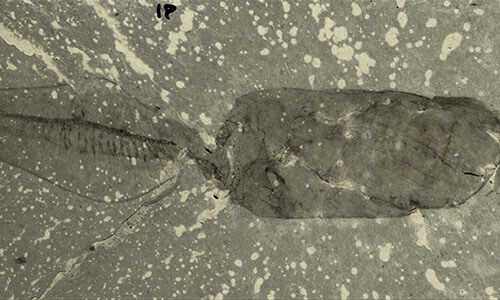
Banffia could be a stem-group hemichordate, a stem-group echinoderm, or a stem-group ambulacrarian. (Ambulacraria is the clade comprising echinoderms (starfish, crinoids and relatives) and hemichordates.)
Another problematic Cambrian fossil: Rotadiscus has a robust upper surface that has some similarity to the urochordate tunic. It lacks a brain, skull, stereom, three-part body, notochord or tail; and has gill slits and feeding appendages. Its other attributes are uncertain.

If the tunic is homologous with that of urochordates, Rotadiscus would be a member of that total group. We would have to account for the absence of a brain, notochord and tail – unless, as with modern urochordates, these three characters were present in larval stages and lost in the fossilized adults. We'd also have to interpret the feeding appendages as convergent traits, leaving a total of four extra steps on our tree.
It is more parsimonious to treat the potential tunic as a homoplasy (convergently derived character), in which case the other characters combine to position Rotadiscus in the stem lineage of Hemichordates and/or echinoderms.
Nectocaris is another Burgess Shale fossil; it has been interpreted as having a muscular tail, but lacks a notochord.

If hearts, brains and notochords are quick to decay, their absence could reflect non-preservation rather than an original biological absence.
If the notochord is genuinely absent, the affinity of Nectocaris can be constrained to quite a precise position in the chordate stem lineage – and Nectocaris would help to constrain the sequence of evolution (i.e. a muscular tail preceded the evolution of the notochord).
However, if the absence of these characters is taphonomic, there is much more uncertainty about its position: it could also sit higher in the stem lineage, or in the chordate crown group, with an affiliation to one or more chordate phyla.
(In fact, the interpretation of Nectocaris has since been substantially revised, and it is no longer thought to be a deuterostome.)



