


Propose and vote for questions or topics to cover during the face-to-face session.
Systematic Palaeontology
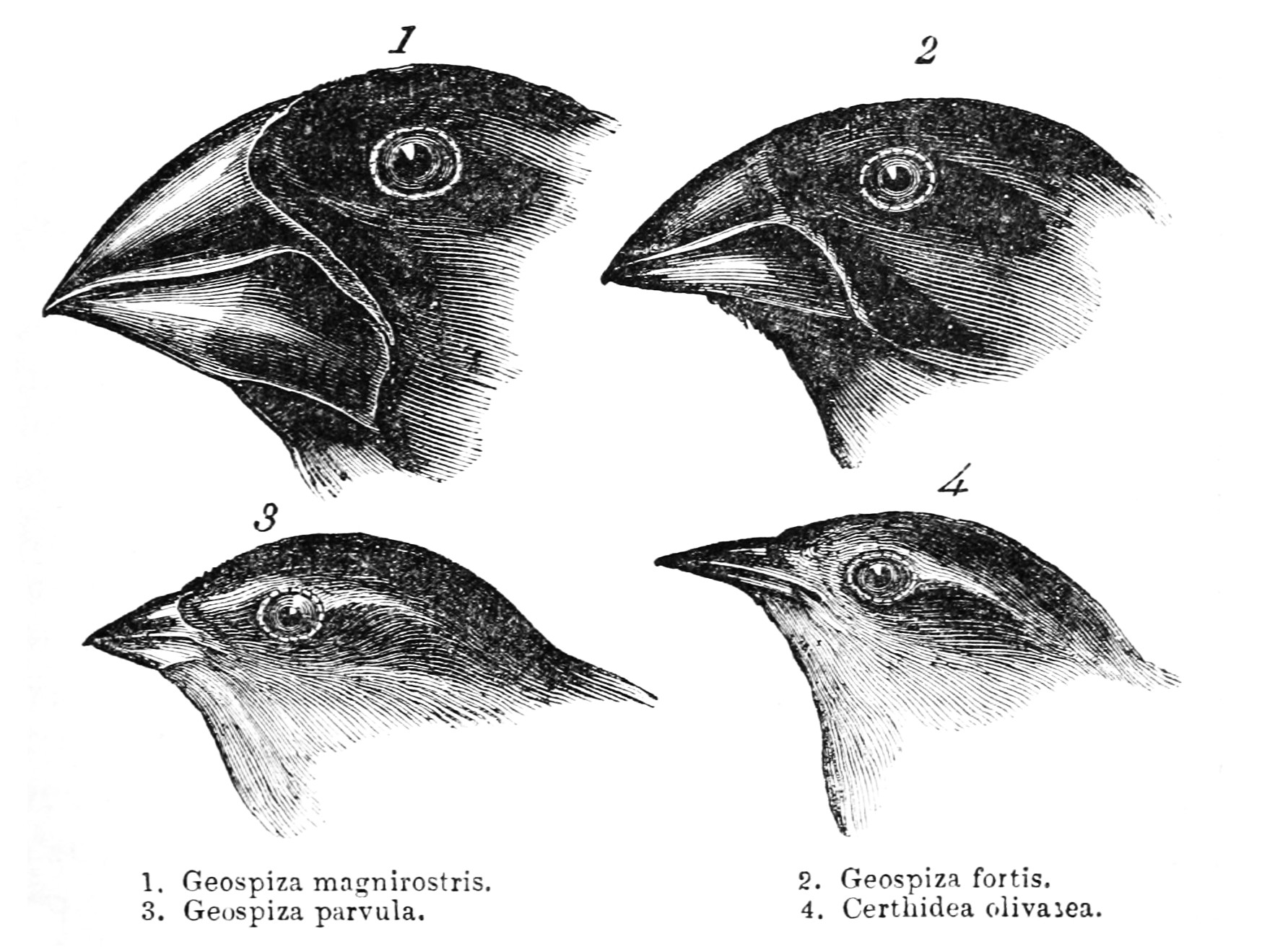
If we want to understand the diversity and complexity of ancient ecosystems, we need to agree what we’re measuring. The number and disposition of species is a good place to start: but what exactly is a species, and why does a fossil belong to Species X and not Species Y?
-
This session introduces lots of new concepts.
You might want to have a separate sheet of paper or browser window to hand
in which to note down brief definitions of
key terms as they are introduced.
Understand how species arise through evolutionary processes
Evaluate different approaches to demarcating species
Appreciate the challenges inherent in recognizing species