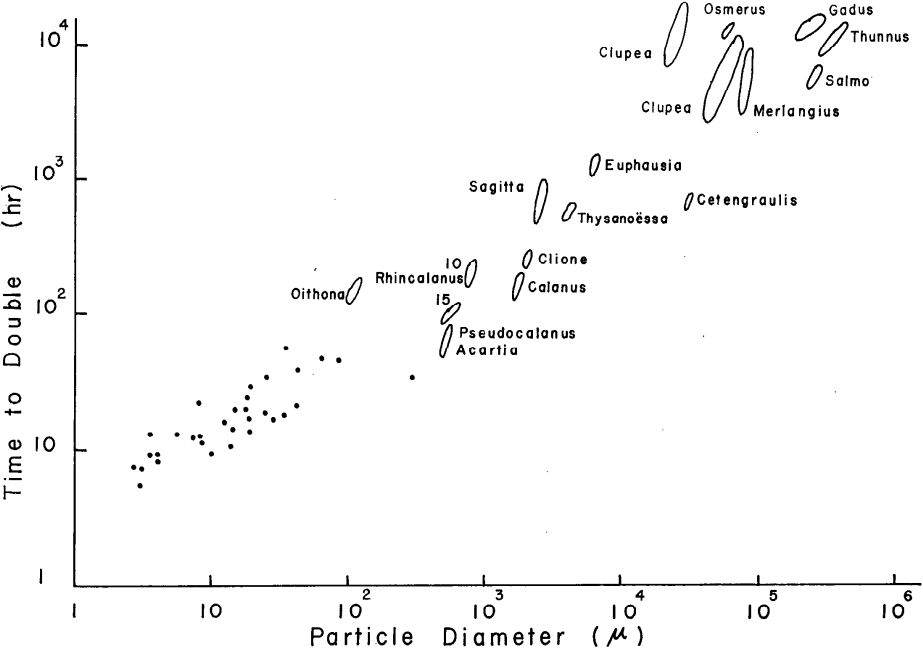
These lifestyles can incidentally complicate the ‘easy’ lives of more efficient organisms.
Interestingly, lineages do not tend to change their ‘energeticness’ / efficiency / effectiveness of nutrient capture very much as they evolve. Rather, new strategies for effective resource use evolve suddenly as a ‘new major group’ develops in a new trophic guild (which came first is an interesting question).
Indeed, lineages seem to show a trend to decreasing evolvability through time: all the ‘low hanging’ evolutionary fruit have been picked, and subsequent mutations are more likely to have a smaller effect. (cf. ‘key innovation’ model of evolution.)
As such, changes in energetics of organisms tend to represent the arrival of new, ‘caffeinated’ lineages (‘bullies’), which displace existing lineages into oligotrophic refugia.
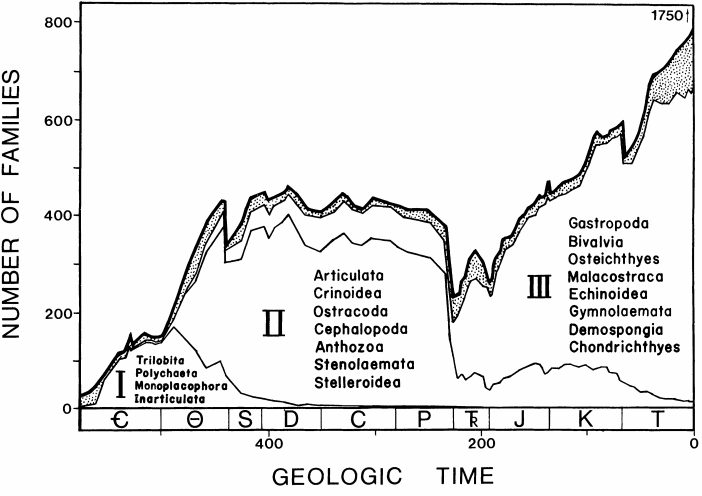
These new lineages are often members of the Recent Fauna (III),
which predominantly comprise high-energy taxa; in the Palaeozoic, the
high energy requirements of such taxa could only be met in the most
productive places on Earth, but the across-the-board increase in
productivity through the Recent meant that their high demands could be
met in progressively more settings – whereas the low nutrient settings
preferred by the Palaeozoic fauna were now found in settings that were
previously too nutrient-scarce to support life at all.
This fits with the long-term picture: diversity is predominantly added into previously unoccupied guilds. Perhaps these were unoccupied because they were not energetically feasible, as not enough biomass was available to sustain them?